CAN CONTACT BINARY THEORY EXPLAIN THE HEAD-TIP AND FRACTURES ON THE ROTATION PLANE? (To be read in conjunction with Part 10).

“The Collisional Problem”. I might have dreamt that up as an apt subtitle for this post because by the time we’re done it will surely be merited but, too bad, I was beaten to it. Those words were already taken as the subtitle for a poster accompanying a talk on 67P’s status as a contact binary. The venue was the American Geophysical Union (AGU) Fall Meeting in December 2014 and the verdict on that binary status remains equivocal.
The abstract for the presentation was entitled,”The Nucleus of Comet 67P/Churyumov-Gerasimenko : a New Case of Contact Binary?”
https://agu.confex.com/agu/fm14/webprogrampreliminary/Paper13889.html
It acknowledged that:
“A contact-binary among the Jupiter family comets (JFC) such as 67P would have profound implications since it must be primordial and the comet must have survived a possible history of collisions in the Kuiper belt. The present cumulative distribution function of size of nuclei of JFC comets indeed suggests a collisionally-relaxed population.”
In other words, there is countervailing evidence that militates against the existence of contact binaries in such a “collisionaly relaxed” population.
The collisional problem is, however, just the first of many for the CB theorists. They might be able to attempt an explanation of the rotation plane head-tip and fractures but not without jumping through flaming hoops in the process. They seem up to the task though. The AGU abstract considered the “unlikely…scenario of a re-accumulated body following a catastrophic collision” (because it really ought to have been blown to smithereens by now), as if it was a last resort to explain the two-lobed shape as a contact binary.
True to say, everything should be considered. Asymmetrical erosion (carving out the lobes via lopsided outgassing) was given a fair hearing too. But it seems the notion of ditching both theories altogether and contemplating stretch theory was just one step too far.
At least one of those authors presented again two weeks later, at the American Astronomical Society’s 225th meeting (AAS 225, 4th to 8th January 2015), still referring to 67P as a contact binary or possibly a single, eroded body. Again, the phrase “a new case of contact binary?” was used, along with the poster, still reminding us of the collisional problem.
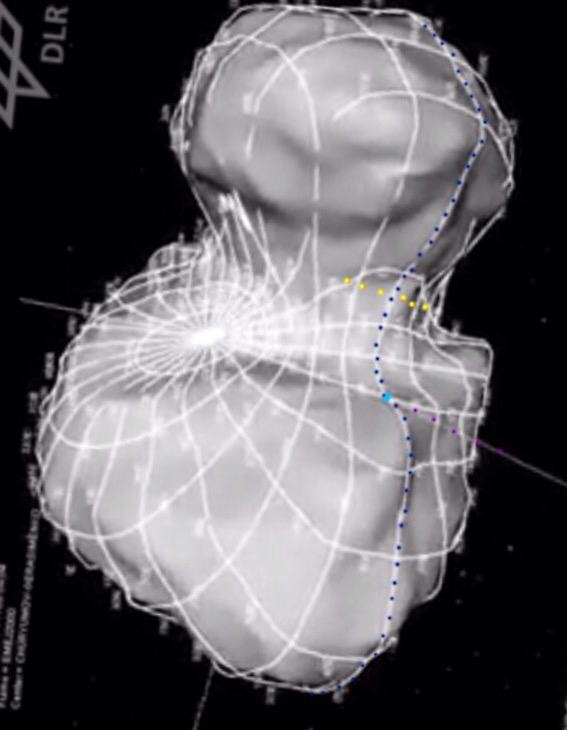
But the answer to the collisional problem was on the poster itself! It had a picture of the comet, looking straight down the z axis of rotation. The visible perimeter, the horizon, was therefore the xy rotation plane and the lower-right horizon was where the fractures happen to straddle the neck at 90 degrees. Furthermore, the uppermost tipping of the head kisses that horizon line too, right above the fractures.
That poster was what made me twig the relationship between rotation plane, spin-up, head tip and fractures that lead to this twinned pair of posts. So the answer to the collisional problem is what it was telling us all along- that 67P can’t be a contact binary. But you had to let go of CB theory fully, if only momentarily so, in order to see the hallmarks of stretch theory depicted on that poster. They are crying out at us: rotation plane, head-tip, fractures.
Here’s a tweet of “the collisional problem” poster:
https://mobile.twitter.com/allplanets/status/545637097797730304
Not much AAS 225 or AGU 14 material is available so we had to rely on tweets. I believe that at least two tweeters quoted here were present at the talks and the third, possibly so, or an astronomer with a live feed of some sort. I can’t be certain of this but they all seemed informed and genuine.
Respondents to the above tweet and elsewhere on the #AGU14 hashtag, some from respected institutions, were happy to support CB theory. One suggested tidal friction leading to tidal locking (of head and body) and then settling against each other. Another supported the asymmetrical erosion tack but no one considered stretching.
Incidentally, I can’t imagine a secondary with one side as flat as that shown in the photo below, offering that side up to the primary in an attempt to lock tidally. It would be inherently unstable. If it ever locked at all it would only be stable if it was the other way round with the topmost ‘crater’ facing into the neck. The reason it’s stable in stretch theory is because it always had a supporting neck to attenuate excessive tipping via compressive resistance to any downward vector on any side. This would even have been the case when it was ‘weightless’ during the stretch because the ‘locking’ referred to above is in two of the three rotational axes of the head, not its translational stretch. There are signs this compressive resistance did indeed happen because the head tip only went so far. Here’s the photo:
 Photo from Part 10. Blue dots: xy rotation plane; yellow dots: ends of one fracture among several.
Photo from Part 10. Blue dots: xy rotation plane; yellow dots: ends of one fracture among several.
Of course, this remarkably flat underside of the head can now be seen for what it is. It’s simply the extension of the now well-documented cliff, reemerging on the other side of the neck. This was the first photo to come out that showed it, proving the head lobe has a flat underside all the way across and overhangs the neck evenly on both sides. The entire plane seats itself back down neatly all round the currently visible portion of the comet body and will doubtless prove to do so on the dark side when it comes into view.
At another AAS 225 presentation, 67P was presented as a contact binary, plain and simple. The presenter pointed out that while several other comets had rocks joined end-to-end, this one had one on top of the other so it looked like a sphinx. That’s actually a far from fatuous distinction, though I’m not so sure anyone was aware of the fact. It should immediately point to the reason the head is tipped up at the back and lead on to the underlying mechanism at play: spin-up, leading to head-tip, stretch and transverse fractures. The head was thrown forward during spin-up due to the very fact that it’s “on top” and not at the axis extremity. But this fact becomes clear only if the rotation plane is considered.
So, yes it does look like a sphinx but the contact binary assumption made on the way to that conclusion is troubling. The “on top” reference was unwittingly portentous but it became just an interesting observation in the absence of any willingness to loosen the grip on CB theory. Here are the relevant tweets:
https://mobile.twitter.com/girlandkat/status/552252787837648896
The presenter went on to say that the fractures in the neck were due to the head rocking against the neck. That’s true up to a point. It’s due to the head tipping forward during the stretch, among other things, but “rocking against” (the tweeter’s words) suggests random movement with no known cause. Here’s the tweet:
https://mobile.twitter.com/lauravican/status/552253732109307906
As if this succession of respected presentations wasn’t enough to leave stretch theory buried for good another American Geophysical Union presentation, on December 18th 2015, managed to take all the evidence as presented in Part 10, which proves the comet stretched and use it, in error, to prove the exact opposite: that 67P’s shape resulted from the head lobe crashing in from the opposite direction. A sure case of a contact binary. Here’s an article from Wired magazine that reported on that presentation:
http://www.wired.com/2014/12/rosettas-comet-shaped-like-duck/
The reasoning went that, seeing as the strata in the head lobe and the body lobe don’t align, they can’t be related and so they must be two different bodies that drifted together. But the only reason they don’t align is the simple fact that the head has tipped up, taking them out of alignment. The Wired article explained that using new images taken with Rosetta’s OSIRIS camera, the presenter and his colleagues found “terrace-like layers” on the comet’s body and that:
“The layering aligns perfectly with parallel lines seen on the opposite side of the body, suggesting that these layers extend through the body as part of its internal structure. Although the head also has layers, they don’t align with those in the body, which implies that the two lobes were once two separate pieces. If the head and body were made from one piece, the layers should extend through both lobes in the same direction.”
This line of thinking presupposes that the only alternative to contact binary theory is asymmetrical erosion gouging a scoop out of a single rock. The only way the presenter could envisage the strata lining up through both lobes was in the scenario where a single body started out with its strata intact all the way across and then this scoop was removed, leaving the two separate lobes but with their strata still in alignment.
That assumption is what steered the presenter away from the most obvious solution- that the head had tipped up, taking the strata out of alignment. If it was tipped back down by 30-40 degrees and seated onto the body, the strata lines would align perfectly, as demonstrated in part 6 of this series.
But it was the head-tip itself that led him to believe that that strata had always been out of alignment so this crucial piece of evidence that so strongly points to stretch theory was used unwittingly and in error to ‘prove’ instead that contact binary theory is correct.
That is a perfect example of the adherence to a cherished theory blocking out even the very thought processes that might lead to the correct theory- a case of fitting new data to an old way of thinking, rather than letting new information speak for itself.
It is also of note that this hypothesing around the finer points of CB theory is based on the highest resolution photos from the OSIRIS camera on the Rosetta orbiter. Just a few scientists are privy to this data at the time of writing (including those with the collisional problem) and it was my honest opinion that they could not help but alight on stretch theory with the abundance of evidence before them, much of it still to come in the next few parts of this series.
However, the Wired article went still further:
“The neck of the comet also shows signs of a collision between the head and the body. The region is covered in big fractures, [the ones straddling the xy rotation plane in the photo above] which would have been created by shockwaves that blasted through the comet during a crash. Some of the fractures are also misaligned, suggesting that they belonged to what were separate, smaller chunks that were floating around when the head merged with the body.”
I have never seen a compressive force cause fractures in a concave surface without it resulting in an explosive shearing event. There’s simply nowhere else for the material to go. Witness compression-testing of concrete pillars that aren’t even concave. They explode dramatically. If further compression of the comet’s neck material is invoked to counter that claim, it is self-contradictory because if it can compress still further it won’t fracture. Indeed, with a porosity of 74%, the neck would have plenty of give in compression and at most it would simply bulge out in folds. But it would not fracture under compression unless it sheared violently as well.
The fractures have clearly resulted from tensile, flexion and torsional forces none of which are compressive (except where flexion compresses the opposite side of the neck).
Incidentally, the mention that the fractures are ‘misaligned’ is interesting. Their average direction is distinctly at right angles to the rotation plane and in successive parallel lines. Yes, they do make noticeable excursions from that simple large-scale picture, presumably due to structural anomalies, but the overall impression is as presented in the photo above and the others in Part 10 (reproduced below).
So, according to this second AGU presentation, as well as the head rocking against the neck (on the sphinx), it had first of all collided with the neck, which was apparently ready-formed, protruding into empty space and pointing in exactly the right direction to make the catch. Once captured, the head ended up perfectly centred with its flat plane conveniently facing downwards, allowing it to overhanging evenly all the way round the neck. That’s quite an impressive claim, provoking at least four tough questions.
Moreover, even if the idea of the colliding head causing the fractures appeared to have some merit at first glance, it wouldn’t explain why those fractures are clustered in parallel lines, at one end of the comet, exactly straddling the rotation plane, at 90 degrees to it, sitting right under the most tipped-up part of the head, and found nowhere else on the neck.
That’s six more really tough questions, ten in total, for the CB adherents to answer regarding the collision of head and body- and that’s the real collisonal problem here, the one that truly merits the subtitle.
The simple answer is that this is where the neck stretched the most and the head tipped forward. Stretch theory would not only answer those 10 questions with ease, it would also predict all ten outcomes as being highly likely.
If they did somehow manage to jump through all ten hoops, the CB theorists would then have to go on to explain why the plan-view matches between head and body were irrelevant- along with their corroborating 3D matches (Parts 1-5). Then explain away the ridges that straddle head and body, followed by the matching strata layers (Part 6). After that, the 30-metre uplifted ‘gull wings’ and slurry piles arising, apparently, from gentle sublimation (Part 7). Then the dykes (Part 8), and the missing slabs (Part 9) -of which there are several more to come- and at least three more as yet unpublished pieces of compelling evidence. That’s eleven more hoops, twenty-one in total to date and counting.
Sorry, twenty-two, I forgot the the original “collisional problem”.
In conclusion, stretch theory answers a multitude of questions that contact binary theory cannot hope to address. Yet on January 8th 2015, the day that AAS 225 closed for business, and astronomers and reporters tweeted views of Seattle en route to the airport, stretch theory had yet to see the light of day.
Copyright ESA/Rosetta/NAVCAM – CC BY-SA IGO 3.0
Here are the other two photos from part 10: